




أهم نظم زراعة الأسطح المكثفة
د. وليد فؤاد أبو بطة
الكاتب : أ. بسام مرستاني
12:20 صباحًا
18, أغسطس 2015
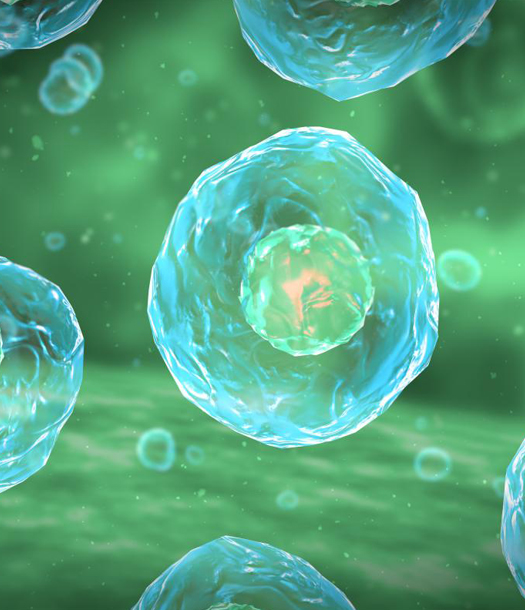
تأتي أهمية دراسة الطبيعة والخواص الكهربائية للأغشية الحيوية المحيطة بالخلايا، وبغض النظر عن تعقيدات العمليات البيولوجية والأجهزة البنيوية والعمليات الأيضية الموجودة في الخلايا والبنية التركيبية للغشاء والمكونة من طبقتين شحميتين فوسفوريتين ذات طبيعة عازلة، وذلك في تحديد الإشارات الكهربائية والناقلية الكهربائية المنتشرة على طول الغشاء الخلوي. كما أنها تلعب دوراً كبيراً في تحديد آليات تحرير الدواء وذلك من أجل معالجة الكثير من الأمراض العضال مثل سرطان الدماغ والألزهايمر والباركسونية …الخ من هذه الأمراض الخطرة.
تكمن صعوبة الوصول إلى دواء من أجل معالجة هذه الأمراض بسبب وجود حاجز (دماغ – دم) الذي يشكل عائقاً أمام وصول الدواء للمنطقة المحددة في داخل الدماغ على سبيل المثال لا الحصر. وبما أن الناقلية الكهربائية تكون محكومة من قبل العديد من البروتينات الموجودة في البنية التركيبية للخلايا والغشاء الخلوي والتي تلعب دوراً كبيراً في تحديد قيمة المقاومة الكهربائية الكلية للخلية والغشاء الخلوي والتي تتكون من أربع مقاومات رئيسية: الأولى عبارة عن مقاومة كبيرة موجودة في الغشاء الخلوي (تتألف من قسمين علوية و سفلية) والتي يمكن اختزالها إلى مكثفة تكون ممثلة لها. والمقاومة الثانية هي مقاومة الوسط الموجود في داخل الخلية.
أما المقاومة الثالثة والتي تسمى بالوصلة المحكمة (Tight junction) والتي تربط الخلايا المتاخمة مع بعضها البعض والمقاومة الرابعة والأخيرة هي مقاومة الفراغ بين الخلايا المتجاورة. وحيث تلعب ما يسمى بقنوات ضخ الأيونات والمسامات وبالتحديد قنوات نقل الصوديوم والبوتاسيوم في الخلية الدور الرئيسي في المقاومة الداخلية والمقاومة الغشائية للخلية سامحة لها بالعبورمن وإلى داخل وخارج الخلية حيث تحول الغشاء إلى مادة ناقلة. أما بالنسبة لمقاومة الوصلة المحكمة تلعب بعض البروتينات الداخلة في تركيبه الدور الرئيسي في تحديد قيمتها بالإضافة إلى عوامل أخرى. وقد وضعت العديد من النماذج الرياضية الهامّة والتي كانت تمثل طبيعة الناقلية الكهربائية في الخلية وغشائها وبالتحديد الخلايا العصبية (العصبونات) والمقاومة الكهربائية عبر الوصلة المحكمة. كما تم تشبيه الغشاء الخلوي على أنه عبارة وصلة نصف ناقلة من نوع PN.
مقدمة عامة
تحاط الخلية بالغشاء الخلوي الذي يحفظ جميع مكوناتها الداخلية حيث يعمل الغشاء البلازمي كحاجز شبه نفوذي بين السطح الداخلي والخارجي للخلية ويمكن تشبيه خارج الخلية بأنها حمام ملحي (ماء مع أيونات منحلة فيه). وفي درجة الحرارة الفيزيولوجية يكون الغشاء الخليوي في الحالة الفيزيائية المائعية و في درجات الحرارة الأبرد يمتلك بنية شبيهة بالبنية الجيلية. حيث يعرف الغشاء البلازميplasma membrane بأنه غشاء حيوي أو غشاء خارجي للخلية مكوناً من طبقتين شحميتين فوسفوريتين (phospholipids) بالإضافة إلى البروتينات.
يشكل هذا الغشاء طبقة رقيقة ونصف نفوذة والتي تحيط بالسيتوبلازما والمكونات الأخرى في الخلية. حيث أن الوحدة الأساسية للحياة البيولوجية هي الخلية والتي تعتبر كتلة من الجزيئات الحيوية في المحاليل المائية (الماء مع الأيونات المنحلة فيها) وتكون محاطة بواسطة الغشاء الخلوي. واحدة من المظاهر المميزة للخلية الحية أنها تضبط تبادل الأيونات المشحونة كهربائياً عبر غشاء الخلية. و تتميز الموائع في خارج الخلية ECFs بأنها متشابهة ماعدا المحتوى البروتيني العالي للبلازما، حيث يكون الصوديوم الكاتيون الأساسي والكلور الأنيون الأساسي بينما تتميز الموائع داخل الخلية بأنها تحتوي على عدد قليل من أيونات الصوديوم والكلوريد. و يكون البوتاسيوم الكاتيون الرئيسي بينما الفوسفات الأنيون الرئيسي.
وبما أن الطبقة الثنائية الدهنية للغشاء البلازمي تشكل حاجزاً عازلاً فاصلاً للمحاليل الملحية الناقلة للكهرباء ما بين داخل وخارج الخلية، فإن الغشاء البلازمي يتصرف كمكثفة، حيث أن سعة المكثفة تعتمد على خواص المادة العازلة بين الصفيحتين والتي تمثل المادة الدهنية في الغشاء البلازمي للخلايا. وتقدر سعة هذ المكثفة تقريباً بحوالي1×10-6 فاراد وذلك لكل (cm2) من سطح الغشاء. وتقدر سماكة الغشاء المحسوبة من خلال العلاقات الرياضية بحوالي (4.5 nm) أو (7.5nm) وذلك حسب التقديرات الميكروسكوبية الإلكترونية. وبالتالي من خلال هذه المقالة سوف نقوم بعرض أهم السمات الكهربائية للغشاء الخلوي، ثم دراسة لآليات الاستقطاب في الغشاء وبشكل مفصل، وأنواع الكمونات التي يخضع لها الغشاء الخلوي، بالإضافة إلى دراسة الطبيعة الكهربائية للوصلة المحكمة (TJ) لما لها من تأثير كبير على قيمة مقاومة الغشاء الخلوي. ثم أهم النماذج الرياضية المكممة من أجل فهم كمون العمل وأخيراً شرح بسيط لنظرية الكوابل لما لها من أهمية في فهم الطبيعة الكهربائية للأغشية الخلوية العصبونية من حيث مفهوم الناقلية السلبية.
الكمون الكهربائي للغشاء
يعرف كمون الغشاء حسب القاموس (Biology –online Dictionary) بأنه النسبة بين التراكيز الداخلية والخارجية لكلاً من أيونات البوتاسيوم، الصوديوم، الكلوريد وأيونات أخرى في الغشاء القابلة للانتشار (diffusible) أو الخلايا. و حسب القاموس الطبي يمثل الفرق في الاستقطاب الكهربائي أو الشحنة بين طرفي الغشاء الخلوي أو جدار الخلية.
أنواع الأيونات الكهربائية الموجودة في داخل و خارج الخلية الحية
تقاد الإشارات الكهربائية ضمن الكائن الحي بشكل عام بواسطة الأيونات. والكاتيونات الأكثر أهمية لكمون العمل هما الصوديوم (Na+) والبوتاسيوم (K+) والكالسيوم (Ca2+). أما الأيونات فهو الكلور ( Cl-) وأيونات أخرى.
ما هو السبب في وجود فرق الكمون بين طرفي الغشاء؟
بكل بساطة كمون الغشاء يعود إلى التفاوت في التركيز والنفوذية للأيونات الهامة عبر الغشاء على سبيل المثال ناقلية الأيون من خلال قنوات أيونية خاصة وبسبب التراكيز الغير متساوية للأيونات عبر طرفي الغشاء فإن الغشاء يمتلك شحنة كهربائية بالإضافة إلى فعالية المضخات الجينية الكهربائية (electrogenic pumps) على سبيل المثال: (Na+/K+-ATPase and Ca++ transport pumps).
السمات و التوصيف الكهربائي للغشاء البلازمي
يعتبر الغشاء الصناعي عازل جيد للكهرباء (لا وجود للأيونات الحرة في داخل الغشاء) وقيمة الناقلية النوعية في واحدة المساحة تكون حوالي ( gpure= 10-13 Ω-1m-2. ويختلف عن الغشاء البيولوجي بأن الأخير يحتوي على أنواع مختلفة من القنوات الأيونية و المسامات والتي تسمح للتيار بالعبورعبر الغشاء رافعة من السلوك المعقد للغشاء ومن قيمة الناقلية الكهربائية بالمقارنة مع الغشاء الصنعي. إن الغشاء بالتعريف يمتلك مكثفة وباعتبار أن الطبقة الدهنية الثنائية تكون رفيعة لدرجة كبيرة وتكون حوالي 6 nm، فإنها لا تحتاج إلى فولتات غشائية كبيرة من أجل خلق حقل كهربائي قوي من خلالها من أجل فصل الشحنات ولذلك فإن سعة الغشاء تكون كبيرة بشكل كافي، وهي تقدر بحوالي (1.10-2 F /m2) في واحدة المساحة. وبشكل غير مشابه للناقلية الموجودة في الغشاء، فإن سعة الغشاء تكون متأثرة بشكل قليل جداَ بتعقيدات العمليات البيولوجية.
دراسة آلية تغيير استقطابية الغشاء الخليوي عند التحفيز
لا بد لنا من دراسة آلية تغيير استقطابية الغشاء الخليوي لما له من تأثير على الطبيعة الكهربائية للغشاء وبالتحديد على ناقلية ومقاومة الغشاء. عند تعرض هذا الغشاء القابل للإثارة مثل الخلايا العصبية والخلايا العضلية لمحفز خارجي (فيزيائي، كهربائي، كيميائي،.. الخ) يسبب في حلقة نزع استقطاب الغشاء، وفرط الاستقطاب ومن ثم العودة إلى قيمة الاستراحة (استقطابية الغشاء في الحالة الطبيعية) والتي تستهلك بحدود ميللي ثانية، ويمكن أن يكون أجزاء بالمئة من الثانية في الخلايا العصبية النموذجية. وهذه التغيرات الدائرة في كمون الغشاء تنتج من الفتح والإغلاق المتعاقب لقنوات الصوديوم في الأول ومن ثم لقنوات البوتاسيوم. و من أجل فهم هذه الآليات لا بد لنا من شرح آليات الاستقطاب التي تحدث في الغشاء وأنواع كمونات الغشاء.
نزع الاستقطاب Depolirization
من حيث المفهوم البيولوجي فهو يشير إلى تغير مفاجئ ضمن الخلية حيث تخضع الخلية من خلالها إلى تغيرات فولتاتية في الكمون الكهربائي للغشاء الخلوي. وهذا يعني بأن داخل الخلية الذي كان سالباً سيصبح موجباً مقارنة مع البيئة الخارجية المحيطة بالخلية. و من أجل تحقيق هذا الغرض تفتح قنوات الأيونات المبوبة كهربائياً والتي كانت مغلقة في حالة كمون الاستراحة. عندما يتم نزع استقطاب الغشاء ستحدث تغيرات تكييفية أو تماكبية conformational في بروتينات القناة والتي تسبب في فتح البوابة عند سطح العصارة الخلوية للمسام وبالتالي فتح المزيد من القنوات الصوديوم ومن أجل أجزاء من الميللي ثانية تصبح نفوذية الغشاء لها أكبر بكثير من نفوذية البوتاسيوم، حيث أن قنوات البوتاسيوم المبوبة كهربائياً تفتح ولكن بشكل خفيف بعد نزع الاستقطاب الأولي وتدعى بقنوات البوتاسيوم المؤخرة delayed بعد ارتفاع كمون العمل. هذا يعني دفع مزيداً من أيونات الصوديوم الموجبة إلى داخل الخلية رافعة من كمون الخلية من السالب إلى الموجب بينما تبقى قناة البوتاسيوم الغير مبوبةnongated التي كانت تعمل في مرحلة الاستراحة بمواصلة العمل. وعندئذٍ تكون عملية نزع الاستقطاب اكتملت.
عملية إعادة الاستقطاب Repolarization
بعد أن تكون الخلية قد نزع استقطابها وخضعت لتغير نهائي في الشحنة الداخلية، و خلال فترة تسمى بفترة التمرد (refractory period) والتي عندها قنوات الصوديوم المحفزة كهربائياً تغلق مرة أخرى بينما تفتح قنوات البوتاسيوم المبوبة كهربائياً بشكل كبير. من أجل القيمة عند قمة كمون العمل فإن الشحنة الموجبة المتزايدة في داخل الخلية تسبب في فتح المزيد من قنوات البوتاسيوم لتتحرك تحت تأثير تخفيض تدرج التركيز نحو خارج الخلية منقصة كمون الخلية ليقترب من كمون الاستراحة مرة أخرى.
فرط الاستقطاب hyperpolarization
تسبب عملية إعادة الاستقطاب في تجاوز قيمة كمون الخلية حيث تبقى أيونات البوتاسيوم بالتدفق نحو خارج الخلية ونحصل على كمون خلية جديد أكثر سلبية من كمون الاستراحة ويكون قريب من كمون أيون البوتاسيوم في حالة التوازن Ek. يعاد تثبيت كمون الاستراحة بواسطة إغلاق القنوات الأيونية المبوبة كهربائياَ.
أنواع الكمونات الكهربائية التي يخضع لها الغشاء الخليوي
كمون الإسترخاء أو الإستراحة Resting Potential: كمون غشاء الاستراحة (PMP) هو عبارة عن فرق الكمون بين طرفي الغشاء عندما تكون الخلية في حالة الراحة. حيث يكون غشاء الخلية أكثر نفوذية لأيونات البوتاسيوم من ايونات الصوديوم وذلك بسبب أن قنوات التسريب (Leak) للبوتاسيوم اكثر من قنوات تسريب الصوديوم. وكما أن مضخات البوتاسيوم و الصوديوم تقوم بضخ كلاً من الأيونين بعكس تدرج تركيزهما، بينما تسمح قنوات التسريب للتحرك عبر الخلية من أجل تخفيض التركيز العالي. مع اتحاد الايونات المضخوخة والمتسربة فإن الخلية تبقى محافظة على كمون غشاء استراحة ثابت. العامل الاساسي المسؤول عن استقرار و توطيد كمون الاستراحة أو الاسترخاء هو القنوات البوتاسية الغير مبوبة وقد عزى العالم (Bernstein) كمون غشاء الاستراحة إلى ثلاث عوامل الأول غشاء الخلية وبشكل انتقائي كان نفوذاً لأيونات البوتاسيوم والثاني تراكيز البوتاسيوم تكون كبيرة في داخل الخلية والثالث أن تراكيز البوتاسيوم في خارج الخلية تكون منخفضة.
بالتالي تنتج القيمة السالبة لكمون الغشاء في حالة الاستراحة من كون نفوذية الغشاء لأيونات البوتاسيوم أكبر بكثير من الأيونات الأخرى في مرحلة كمون الاسترخاء (حيث أن القنوات المبوبة تكون مهملة في هذه المرحلة) حيث يجري البوتاسيوم بسبب التدرج الكيميائي أو تدرج التركيز القوي من داخل عصارة الخلية إلى خارج الخلية حاملاً معه الشحنات الموجبة. إن قنوات الصوديوم المبوبة كهربائياً تكون مغلقة في حالة كمون الاستراحة. إن عمل الخلية الحقيقي يكون أكثر تعقيداً سامحاً للعديد من الأيونات لكي تساهم في كمون الاسترخاء. هذا يعني بأن كمون الاسترخاء يُسيطر عليه بواسطة أنواع عديدة من الأيونات. حيث أن كمون الاسترخاء يحسب باستخدام معادلة جولدمان أو التي تسمى Goldman–Hodgkin–Katz voltage equation.
كمون العمل Action Potential
يتم توليد هذا الكمون بواسطة أنواع مختلفة من القنوات الأيونية المبوبة فولطياً. وعندئذ تفتح هذه القنوات من أجل عكس استقطابية الغشاء وبالتالي عندما تفتح قنوات أيونات الصوديوم المبوبة كهربائياً بشكل كافي فإنها تتغلب على جريان قنوات البوتاسيوم التي كانت من الأساس مفتوحة في حالة الاستراحة مولدة شحنة موجبة في داخل الخلية مقارنة مع خارج الخلية. ونلاحظ أن مقدار كمون الغشاء عند قمة نزع الاستقطاب في كمون العمل يقترب جداً من كمون التوازن للصوديوم ENa والمعطى بمعادلة (Nernst) وبالتالي إن فتح قناة الصوديوم سيكون مسؤول عن توليد كمون العمل. حيث أنه من الناحية الطاقية و التوازن الثرموديناميكي تفضل أيونات الصوديوم الدخول إلى داخل الخلية وذلك بسب تدرج التركيز و كمون الغشاء السالب في الداخل.
هناك نوعان أساسيان من كمونات العمل، الأول عائد لقنوات الصوديوم المبوبة كهربائياً والثانية لقنوات الكالسيوم المبوبة كهربائياً. كما أن كمون العمل الذي يطبق بشكل موضعي على قطعة من الغشاء يكون له القدرة على تحفيز القطع المجاورة بما يشبه الانتشار في قطع الدومينو المفاجئ ومن دون تخفيف، ولكن كيف؟ عند قمة كمون العمل الانتشار السلبي للأغشية المنزوعة الاستقطاب يكون كافي لنزع استقطاب القطاعات المجاورة في الغشاء، مسبباً في فتح بعض قنوات الصويوم الأيونية المبوبة كهربائياً في هذه المنطقة وبالتالي الازدياد في مدى اتساع إزالة الاستقطاب في المنطقة يتسبب في فتح سريع جداً لأكثر القنوات الأيونية ومن ثم توليد كمون العمل. يحفز ازالة الاستقطاب فتح قنوات البوتاسيوم المبوبة كهربائياً و إعادة كمون الاستراحة. و بالتالي فهو ينتشر على شكل موجة مسافرة بعيداً عن موقعها الأصلي الأول من دون تخفيف.
كمون التوازن Equilibrium potentials
يمثل قيمة فولتائية الغشاء و التي عندها تتوازن كلاً من القوتين الأولى تدرج التركيز الكيميائي والتي تسبب في تحريك الأيون من التركيز المرتفع إلى التركيز المنخفض، والثانية قوى التدرج الكهربائية، و عندما يكون هذان المقداران متساويان بالقيمة و متعاكسان بالاتجاه نقول أن الايون في حالة توازن الكتروكيميائي، بالتالي فإن التيار الصافي للأيون عبر الغشاء يكون مساوياً للصف. على سبيل المثال نجد أن أيون البوتاسيوم يتحرك في حالة كمون الاستراحة من الداخل إلى الخارج تحت تأثير تدرج التركيز الكيميائي بسبب أن تركيز البوتاسيوم في الداخل يكون أكبر من خارج الخلية و يتحرك في نفس الوقت نحو داخل الخلية بسبب الفولتائية السالبة في داخل الخلية. وهذا يعني بأن كمون الغشاء يساوي كمون التوازن ويساوي كمون الاستراحة Vm=VREST =VK.[B2] و عندما يوجد أكثر من نوع واحد من الأيونات في داخل الخلية وهي الحالة الحقيقية عندئذ نسمي كمون التوازن بإسم كمون الاستراحة أو كمون التسرب.
تمتلك الوصلة المحكمة توعين من الوظائف كبوابة و كسياج. حيث يمكننا تبيان و اكتشاف عمل البوابة من خلال قياس المقاومة الكهربائية TERع بر الخلية الظهارية transepithelial للأنسجة. حيث تظهر الدارة الكهربائية بأن المقاومة المذكورة سابقاً عبارة عن مقاومتين موصولتين على التوازي، الأولى تمثل المقاومة عبر الخلية و الثانية المقاومة ما بين الخلايا paracellular والمقاومة عبر الخلية وهي عبارة عن مجموع كلاً من المقاومتين القمية والقاعدية للغشاء اللتان تكونان موصولتان على التسلسل. وبما أن قيمة المقاومة عبر الغشاء البلازمي عالية جداً في الخلايا الظهارية حوالي (2 G Ω). لذلك نجد أن التيار يجري من خلال المقاومة الثانية أي (ما بين الخلايا) والمضبوطة من خلال المقاومة المقدمة من الوصلة المحكمة و لهذا فإن قيمة المقاومة (TER) بشكل عملي تمثل و تعكس قيمة مقاومة الوصلة المحكمة.
كما يمكننا التحكم بقيمة المقاومة الكهربائية السابقة من خلال تعريض الحاجز لمحفز خارجي محدد مثل تطبيق قوى أو إجهاد القص على قنوات الحاجز BBB أي حاجز (دم-دماغ) وتحديد الحالة الأمثلية لعمل الحاجز، حيث أن إجهاد القص يمتلك تأثير لتحويل التنبيه الميكانيكي إلى فعالية (كيميائية – كهربائية) وذلك على العديد من الممرات الجزيئية للخلايا الباطنية وذلك بواسطة تنشيط المستقبلات المرتبطة بالغشاء. تتراوح قيمة المقاومة (TEER) بالنسبة لحاجز (دم-دماغ) عبر الخلايا الباطنية (endothelia) ما بين (1500 -2000 ( Ω.Cm2 و هذا يساعد في تنظيم دخول وعبور الجزيئات المنقولة عن طريق الدم إلى الدماغ ويساعد في المحافظة على التوازن الأيوني ضمن البيئة الميكروية الدماغية. وفي دراسات سابقة قام بها benson وزملاؤوه في معهد الكيمياء الحيوية في جامعة Westfälische Wilhelms-Universität Münster من أجل مراقبة وقياس المقاومة الكهريائية (TEER) في الخلايا الباطنية والظاهرية وذلك بتطبيق قانون أوم باستخدام جهاز بسيط مكون من مجموعتين من الإلكترودات والمسماة (Voltohmeters) ومطيافية أو ما يسمى بالتحليل الطيفي للممانعة (impedance spectroscopy) وقد وجد بأنه عند تطبيق تيار متناوب على خلية أحادية الطبقة تتغير قيمة الممانعة الكلية للخلية مع تغير توتر التيار المطبق.
النماذج الرياضية المستخدمة في تمثيل الخواص الكهربائية لبنية الغشاء الخليوي
بسبب أهمية النماذج الرياضية الموضوعة من قبل العديد من العلماء والتي كانت تمثل صورة حقيقة عن عمل و طبيعة كمونات الغشاء الخلوي إلى حد ما، لابد لنا من تقديم فكرة موجزة عن أهم هذه النماذج الرياضية. حيث قام علماء الفيزيولوجيا الكهربائية وذلك بوضع نموذج للعديد من الأغشية الخلوية، و هي تتألف من مكثفة موصولة على التوازي مع بطاريات وهي تمثل ممرات للعديد من أنواع الأيونات، وكل من هذه البطاريات تكون موصولة على التسلسل مع موصلات و التي تسمى أيضاً بالناقلياتconductance أو مقاومات (resistors ) حيث فيزيائياً تعتبر المقاومة مقلوب الموصلية. وتحدد قيمة هذه السعة بواسطة خواص الطبقة الدهنية الثنائية. أما قيمة الموصلات أو المقاومات فتحدد من خلال حالة القنوات الأيونية والتي تكون نفوذة لهذا الأيون، ضامة قنوات التسرب، قنوات المبوبة بواسطة الليغيندات و القنوات الايونية المبوبة بواسطة الكمون الكهربائي. إن النماذج الرياضية و الحاسوبية من الأمور الجوهرية لفهم كمون العمل، و تقدم التنبؤات التي يمكن ان تختبر بالمقارنة مع النتائج التجريبية .
نموذج Hodgkin-Huxley
استطاع العالمان Hodgkin و Huxley أن يضعا نموذج عن فيزيولوجيا المحوار (زائدة طويلة من العصبون تنقل الاشارة العصبية من الخلية) العملاق لحيوان رخوي ( squid ( والذي يعتبر الحجر الأساس في علوم الأنظمة العصبية. وهو من أكثر النماذج أهمية و دقة و الذي يصف كمون العمل بواسطة مجموعة متزاوجة من أربع معادلات تفاضلية و يمكن تبسيط هذا النموذج مقارنة مع الغشاء العصبي الحقيقي كما هو متواجد في الطبيعة.
(Hodgkin–Huxley) وهو توصيف فيزيائي حيوي للغشاء و يتكون من الموصلتان conductances GNa ،GK المتغيرتان مع الزمن والكمون المطبق (الصوديوم و البوتاسيوم) والموصلة الأخيرة التسريب (leak) GLk التي تكون غير تابعة للزمن و كمون الغشاء. وتعبر المكثفة C عن الطبقة الثنائية الدهنية العازلة، و حيث يمثل التدرج الإلكتروكيميائي المحرك والدافع لجريان الأيونات كبطارية (E) من خلال المنابع الفولتية والتي تكون محددة بواسطة النسبة للتراكيز في داخل و خارج الخلية للأنواع الأيونية. حيث أن ENa، EK، VLk تمثل الجهد (فرق الكمون) لبطارية أيون الصوديوم و أيون الكالسيوم، و كمون التسريب بالنسبة لكمون الاستراحة. بينما (IS) يمثل تيار المحفز الخارجي المطبق وذلك من أجل الحصول على كمون العمل. و بالتالي تعطى معادلة تغير كمون الغشاء مع الزمن في حالة كمون العمل بالعلاقة التالية:
نموذج FitzHugh–Nagumo
في منتصف 1950 بحث العالم FitzHugh في تخفيض نموذج Hodgkin – Huxley إلى نموذج ذو متغيرين و التي من خلالها تطبق تحليل مستوي الطور. وملاحظاته العامة كانت بأن المتغيرات المبوبة gating n،h تمتلك حركية بطيئة بالنسبة للمتغير m وهي عبارة عن متغيرات مبوبة مرتبطة بالكمون. وبالتالي هذا يقودنا إلى نموذج المعادلة بمتغيرين، و الذي يسمى بنموذج مستوى الطور (البطيء –السريع).
نظرية كابل في الناقلية السلبية Cable Theory (Passive Conduction)
لابد لنا من ذكر نظرية الكوابل لأهميتها في حساب التيارات الكهربائية وذلك على طول المحوار neurites السلبي، وبشكل عملي التغصنات dendrites والتي تستقبل الإشارات أو المعلومات السينابتيكيsynaptic وذلك في مواقع وأزمنة مختلفة. والكمونات السلبية عبارة عن سلوك هام مترافق مع الفروع المتغصنة والطرفية للعصبون. إن الكمون السينباتكي يتقدم بشكل موهن attenuation والتي دفعت بالعالم Hermann لينسب هذه العلاقة مع النظرية التي تصف خطوط النقل المفقودة lossy.
أهم المبادئ التي تعتمد عليها النظرية
الخاتمة
لقد وجدنا من خلال الدراسة السابقة كيف أن التحكم بالخواص البنيوية (البروتينات المكونة للغشاء الخلوي)، والخواص الفيزيائية للوصلة المحكمة TJ يلعب دور كبير في تحديد مقاومة الغشاء البلازمي بالإضافة إلى القنوات أو البوابات البروتينية والتي تسمح بعبور نوع محدد من الأيونات الكهربائية أو الجزيئات وفق آليات مختلفة مثل النقل السلبي، الانتشار البسيط، التناضح، الانتشار المحسن، والفلترة، النقل الفعال، الحويصلة والتي تلعب أيضاً دور كبير في تحديد الخواص الكهربائية للغشاء وعملية نقل الاشارات بالإضافة الدور الأساسي في تحديد نوعية تحرير الدواء إلى منطقة محددة من الدماغ على سبيل المثال لا الحصر. في دراسات سابقة تم تشبيه الغشاء الخلوي الوعاء الشعري للعصب بنموذج الوصلة الكهربائية المعرفة PN وحركة حاملات الشحنة في الغشاء (الصوديوم و البوتاسيوم) ما بين داخل وخارج الخلية بحركة الإلكترونات والثقوب في طرفي الوصلة PN.
وهذا التشبيه يعود بنا الى علم محاكاة الطبيعة ذلك العلم الرائع والذي اتحدى به كل العلوم الأخرى بأنه سيكون المفتاح و اللاعب الأساسي في حل الكثير من المشاكل العلمية و الطبية وأولها إيجاد الدواء للعديد من الأمراض. حيث أن كل الأمور والتطورات التكنولوجية في العلوم المتقدمة إنما تعود بأصولها إلى الطبيعة الخلاقة و المبدعة. وأهمية هذه النماذج الرياضية التي ساهمت كثيراً في توضيح المفاهيم و المكونات الكهربائية للخلية الحية و الغشاء الحيوي والعصبونات (المحوار) بشكل خاص. وإن إمكانية وضع نماذج جديدة تضيف قفزة نوعية على مقدار فهمنا للآليات الكهربائية وانتقال الأيونات الكهربائية عبر الغشاء في داخل الخلية وخارجها التي بها من التعقيد في العمليات البيوبوجية ما يكفي لعدم قدرتنا على استيعابها وفهم آلياتها بشكل جيد.
المراجع
البريد الإلكتروني للكاتب: bassam7541@hotmail.com
الكلمات المفتاحية





الموقع قائم على مبدأ الترخيص العام للجمهور في حرية النسخ والنقل والاقتباس من جميع المحتويات والنشرات والكتب والمقالات، دون مقابل وبشكل مجاني أبدي، شريطة أن يكون العمل المستفيد من النسخ أو النقل أو الاقتباس متاحا بترخيص مجاني وبذات شروط هذا الموقع، وأن تتم الاشارة إلى منشورنا وفق الأصول العلمية، ذكرا للكاتب والعنوان والموقع والتاريخ.
هذا والموقع يساعد المؤلف على نشر إنتاجه بلا مقابل من منفعة معنوية أو مادية، شريطة أن يكون العمل متوفراً للنسخ أو النقل أو الاقتباس للجمهور بشكل مجاني. ثم إن التكاليف التي يتكبدها الموقع والعاملون عليه تأتي من مساعدات ومعونات يقبلها الموقع ما لم تكن مرتبطة بأي شرط مقابل تلك المعونات.


د. وليد فؤاد أبو بطة

الكاتب : عبد الحكيم محمود
الكاتب : الصغير الغربي

الكاتب : المحرر

الكاتب : المحرر
الكاتب : أ.د. عبدالرؤوف علي المناعمة

الكاتب : الحسين بشوظ

الكاتب : بيان القاضي
الكاتب : د. طارق قابيل

الكاتب : محمد عارف






يسعدنا أن تشاركونا أرائكم وتعليقاتكم حول هذهِ المقالة عبر التعليقات المباشرة بالأسفل أو عبر وسائل التواصل الإجتماعي الخاصة بالمنظمة
اشترك في نشرات أرسكو ليصلك كل جديد
